

機能詳細
タンパク質/核酸/ペプチドモデリング
 タンパク質の立体構造予測(ホモロジーモデリング)
タンパク質の立体構造予測(ホモロジーモデリング)
ホモロジーモデリングを用いて、アミノ酸配列からタンパク質の立体構造を予測します。ホモロジーモデリングの各ステップをローカルPC内で行えます。抗体専用のモデリング機能や、変異体モデリング機能も搭載しています。
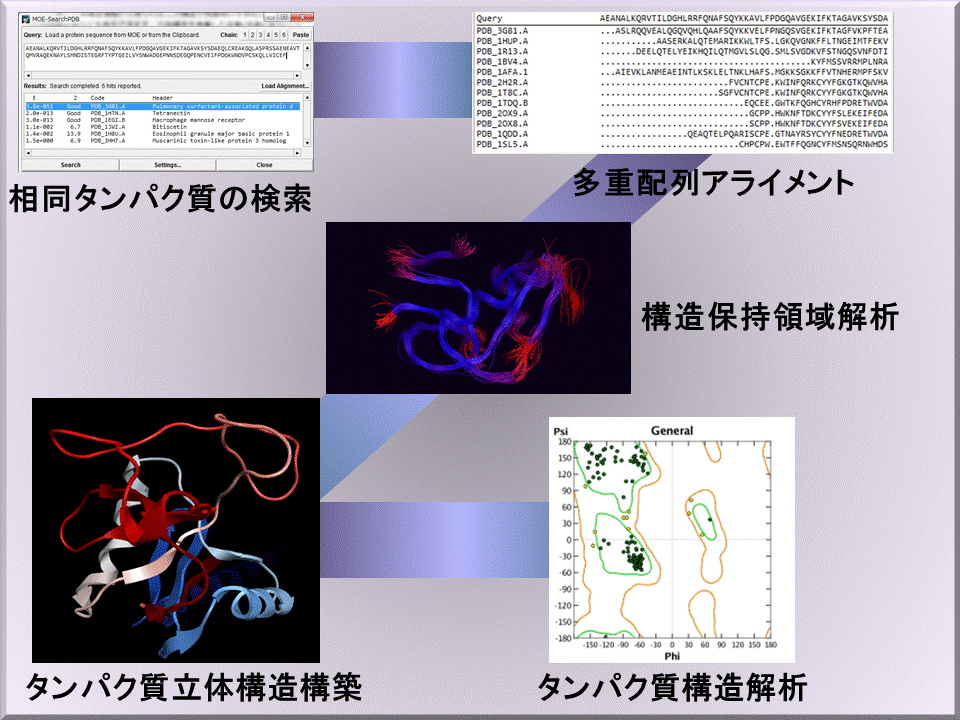
ホモロジーモデリングでは、構造既知である相同タンパク質の構造保存領域の情報を元に、構造未知のタンパク質立体構造を予測します。高精度のモデリングを行う過程として、(1)相同タンパク質の検索、(2)多重配列アラインメント、(3)構造保持領域/相互作用解析、(4)タンパク質立体構造構築、(5)タンパク質構造解析というステップで、類縁構造の検出から相互作用を考慮したアラインメントの補正、得られたモデルの評価を行えます。MOEでは単量体だけでなく、多量体の構造予測もできます。タンパク質立体構造構築の際に、リガンドや水分子、金属イオンなどのヘテロ原子を考慮できます。複数のタンパク質をテンプレートとしたモデリングも可能です。
抗体モデリング
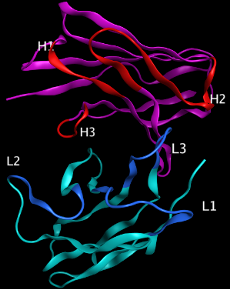 MOEは簡単な操作で適切な抗体モデリングを行うことができます。抗体配列を入力することで、自動的に相補性決定領域(CDR)とフレームワーク領域を認識して領域ごとにテンプレート構造の検索を行い、抗体のホモロジーモデリングを行います。それぞれの領域について配列類似性と構造のバリエーションを考慮したテンプレート候補が提案されます。構築範囲として、Fv、Fab、F(ab’)、rIgG、IgG、VHH、VLL、二重特異性抗体(bispecific抗体)を指定できます。テンプレート構造として整理された抗体データベースには in-houseデータを追加することができ、公共データと in-houseデータを一括して検索することが可能です。複数配列を入力とした抗体構造の連続モデリングも可能です。
MOEは簡単な操作で適切な抗体モデリングを行うことができます。抗体配列を入力することで、自動的に相補性決定領域(CDR)とフレームワーク領域を認識して領域ごとにテンプレート構造の検索を行い、抗体のホモロジーモデリングを行います。それぞれの領域について配列類似性と構造のバリエーションを考慮したテンプレート候補が提案されます。構築範囲として、Fv、Fab、F(ab’)、rIgG、IgG、VHH、VLL、二重特異性抗体(bispecific抗体)を指定できます。テンプレート構造として整理された抗体データベースには in-houseデータを追加することができ、公共データと in-houseデータを一括して検索することが可能です。複数配列を入力とした抗体構造の連続モデリングも可能です。MOE抗体モデリング機能(PDF)
タンパク質ドメイン構造データベースとタンパク質ファミリーデータベース
 PDBデータから構造の冗長性を排除したタンパク質ドメイン構造データベースを用意しています。これはホモロジーモデリングのテンプレートに使用します。また、タンパク質ドメイン構造データベースとUniProtの配列データを用いて、配列および立体構造ベースのクラスタリングにより作成された、タンパク質ファミリーデータベースも用意しています。このデータベースは相同タンパク質の検索対象として利用され、遠縁の構造類似タンパク質の検出やファミリー単位の解析を実現します。
PDBデータから構造の冗長性を排除したタンパク質ドメイン構造データベースを用意しています。これはホモロジーモデリングのテンプレートに使用します。また、タンパク質ドメイン構造データベースとUniProtの配列データを用いて、配列および立体構造ベースのクラスタリングにより作成された、タンパク質ファミリーデータベースも用意しています。このデータベースは相同タンパク質の検索対象として利用され、遠縁の構造類似タンパク質の検出やファミリー単位の解析を実現します。アミノ酸/核酸配列アライメント
MOEは独自の配列と立体構造を考慮したタンパク質/核酸アラインメント手法を搭載しています。この手法は1対1のアラインメントだけでなく、複数のタンパク質/核酸を同時にアラインメントすることもできます。複数の多量体の配列からChainごとにサブユニットを定義することで、サブユニットごとのアラインメントが可能です。配列の一部分や2次構造の重み付け、拘束を考慮したアラインメントが可能です。アラインメントの中で配列のみのデータと構造データを混ぜることによって、構造未知の配列データに対して、信頼性のある推察を得ることができます。核酸では5’-3’あるいは3’-5’の順序を切り替えてアラインメントが可能です。
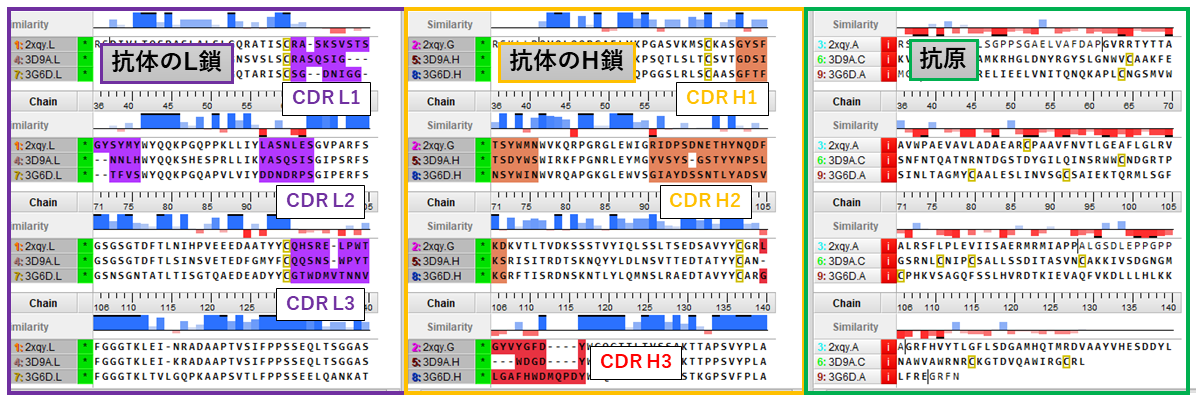 3つの抗体-抗原複合体の配列アラインメント。抗体のL鎖(紫)、H鎖(橙)、抗原(緑)を3つのサブユニットに分けて、それぞれをアラインメントし重ね合わせを実行。CDR L1-3とCDR H1-3は、抗体の抗原認識領域。
3つの抗体-抗原複合体の配列アラインメント。抗体のL鎖(紫)、H鎖(橙)、抗原(緑)を3つのサブユニットに分けて、それぞれをアラインメントし重ね合わせを実行。CDR L1-3とCDR H1-3は、抗体の抗原認識領域。
タンパク質/核酸重ね合わせ
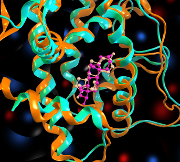 タンパク質/核酸の立体構造の重ね合わせを行い、その結果をレポート形式で出力します。PDB IDなどでタンパク質に関連付けられたリガンドや溶媒などは自動的にタンパク質に追従して重ね合わせられます。また、多量体同士をまとめて重ね合わせたり、受容体ポケット構造など注目している残基セットのみを利用した重ね合わせも行えます。レポートでは重ね合わせ全体のRMSDだけでなく、残基毎のRMSD値がグラフ表示され、構造の異なる残基を容易に検出できます。
タンパク質/核酸の立体構造の重ね合わせを行い、その結果をレポート形式で出力します。PDB IDなどでタンパク質に関連付けられたリガンドや溶媒などは自動的にタンパク質に追従して重ね合わせられます。また、多量体同士をまとめて重ね合わせたり、受容体ポケット構造など注目している残基セットのみを利用した重ね合わせも行えます。レポートでは重ね合わせ全体のRMSDだけでなく、残基毎のRMSD値がグラフ表示され、構造の異なる残基を容易に検出できます。コンセンサス解析
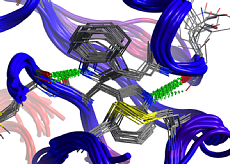 複数のタンパク質に保存されている配列や構造を保持領域として抽出できます。保持領域の定義には残基の一致度、主鎖または側鎖のRMSD、水分子の配置位置が利用可能です。構造保持領域のみに基づく立体構造の重ね合わせや、重ね合わせに基づいた配列アラインメントの最適化、座標平均モデルの構築機能を搭載しています。
複数のタンパク質に保存されている配列や構造を保持領域として抽出できます。保持領域の定義には残基の一致度、主鎖または側鎖のRMSD、水分子の配置位置が利用可能です。構造保持領域のみに基づく立体構造の重ね合わせや、重ね合わせに基づいた配列アラインメントの最適化、座標平均モデルの構築機能を搭載しています。共通する相互作用として水素結合、VDW相互作用、アレーン接触、イオン結合、ジスルフィド結合、配位結合を抽出できます。相互作用ネットワークの認識やアラインメントからの相互作用予測、タンパク質間相互作用の検出も行えます。
タンパク質立体構造の妥当性評価
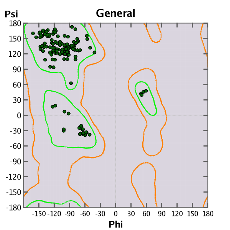 X線結晶解析データから統計的に導かれたデータを元に、モデリングされたタンパク質立体構造の妥当性を評価することができます。Ramachandran PlotやChiPlotプロットなどの二面角のプロット図では、残基毎の各二面角の分布や、異常値を持つ残基をグラフィカルに分かりやすく表示します。また、二面角、結合距離、結合角などの立体化学的な情報をレポートとして出力し、統計的に異常な値を検出します。検出された情報を元に、異常構造を持つ部分にターゲットを絞って構造最適化計算を行うことにより、タンパク質立体構造の修正を簡単に行うことができます。
X線結晶解析データから統計的に導かれたデータを元に、モデリングされたタンパク質立体構造の妥当性を評価することができます。Ramachandran PlotやChiPlotプロットなどの二面角のプロット図では、残基毎の各二面角の分布や、異常値を持つ残基をグラフィカルに分かりやすく表示します。また、二面角、結合距離、結合角などの立体化学的な情報をレポートとして出力し、統計的に異常な値を検出します。検出された情報を元に、異常構造を持つ部分にターゲットを絞って構造最適化計算を行うことにより、タンパク質立体構造の修正を簡単に行うことができます。
ペプチドモデリング
天然アミノ酸あるいは特殊アミノ酸のフラグメントをつなげて簡単にペプチドを構築できます。ペプチドの配列からも構築できます。ペプチドの2次構造として、直鎖、ヘリックス、ストランドなど自由に選べます。天然アミノ酸、特殊アミノ酸への変異も可能で、アミノ酸配座ライブラリーからパッキング、水素結合、二面角などで妥当な側鎖配座を選べます。環状ペプチドの構築も対応し、LowModeMDによる配座解析と組み合わせて妥当な立体構造を出力できます。
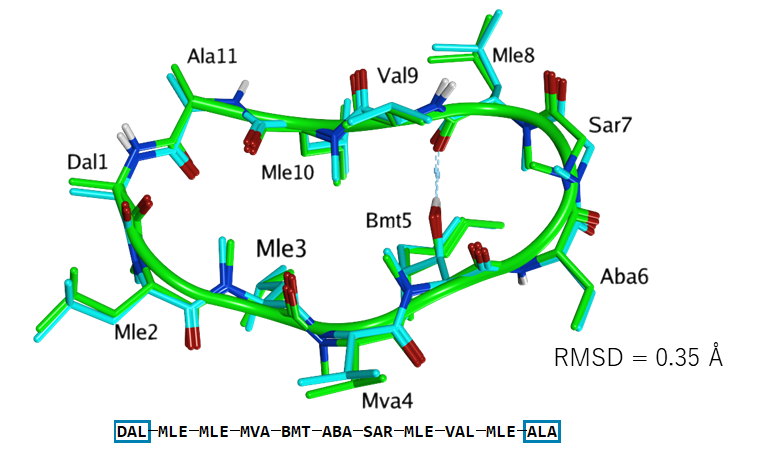
MOEで構築した環状ペプチド(シクロスポリンA)の構造(淡青)とX線構造(緑)との比較。立体配座はLowModeMDで予測。主鎖のRMSD値は0.35 Å。
核酸モデリング
MOEは塩基を順番に選択、あるいは、塩基配列を入力するだけで目的の核酸を構築できます。らせんのタイプとしてA、B、C、Z型を指定した、二重らせん、あるいは、一重らせんの構造を構築できます。一重らせん構造からの相補的な二重らせん構造への補完も可能です。DNAからRNAまたはその逆の変換、指定したらせん構造への変換が可能です。天然塩基への変異体の構築や、特殊塩基を用いた変異体も構築できます。特殊塩基はユーザー定義の塩基を追加できます。
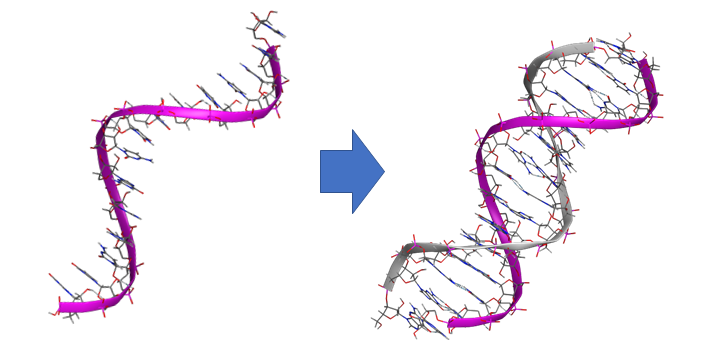
一重らせん構造から二重らせん構造への相補的配列と構造の自動補完。アンチセンス核酸医薬設計を支援。
タンパク質/核酸 変異体モデリング
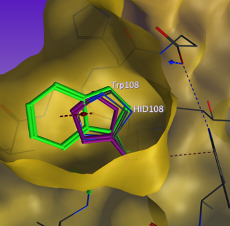 ターゲットとなるアミノ酸残基や塩基を指定するだけで、変異体の配座候補を得ることができます。事前に用意された側鎖配座データベースから、立体障害のない候補構造を提案します。それぞれの候補はファンデルワールスエネルギー、水素結合エネルギーなどによってランク付けされます。特殊アミノ酸、特殊塩基への変異も可能です。
ターゲットとなるアミノ酸残基や塩基を指定するだけで、変異体の配座候補を得ることができます。事前に用意された側鎖配座データベースから、立体障害のない候補構造を提案します。それぞれの候補はファンデルワールスエネルギー、水素結合エネルギーなどによってランク付けされます。特殊アミノ酸、特殊塩基への変異も可能です。
ユーザーオリジナルの特殊アミノ酸、特殊塩基を登録し変異体構築に用いることができます。
タンパク質デザイン/ペプチドデザイン機能
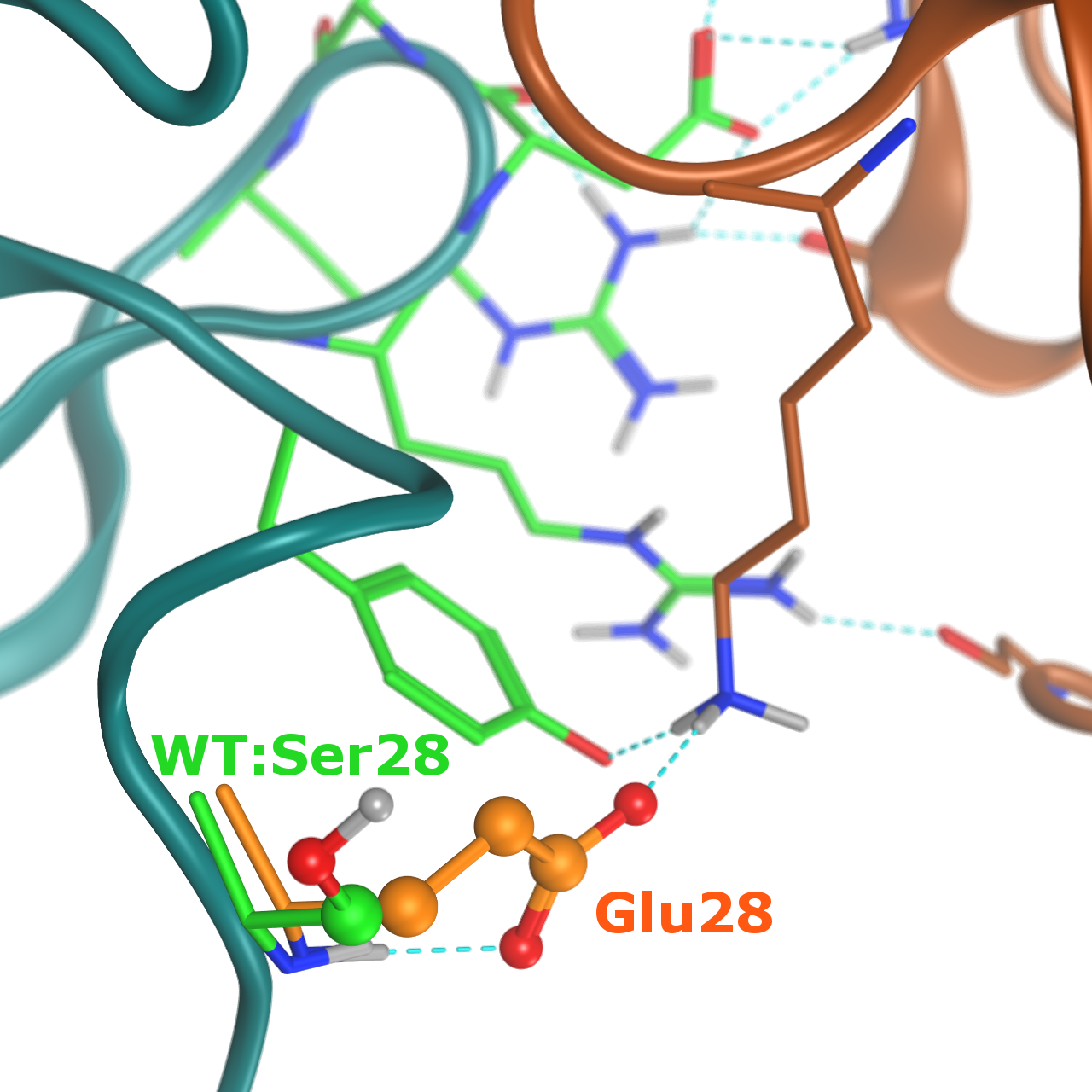 タンパク質の変異体解析をin silico で行うことが出来ます。リガンドとの親和性に重要な残基の解析(ホットスポット解析)や、タンパク質の熱安定性に影響を与える変異体の予測などが行えます。更には変異体の様々な物性値を推算し、野生型の物性値と比較することで、実験の目的に応じた変異体作製に貢献できます。右図は抗体のCDRのセリンを別のアミノ酸残基に変異し抗原との相互作用エネルギーが改善された例です(PDB ID: 1VFB)。グルタミン酸への変異により、抗原のリジンとの水素結合が新たに形成されています。またタンパク質の熱安定性の評価も同時に行うことができます。
タンパク質の変異体解析をin silico で行うことが出来ます。リガンドとの親和性に重要な残基の解析(ホットスポット解析)や、タンパク質の熱安定性に影響を与える変異体の予測などが行えます。更には変異体の様々な物性値を推算し、野生型の物性値と比較することで、実験の目的に応じた変異体作製に貢献できます。右図は抗体のCDRのセリンを別のアミノ酸残基に変異し抗原との相互作用エネルギーが改善された例です(PDB ID: 1VFB)。グルタミン酸への変異により、抗原のリジンとの水素結合が新たに形成されています。またタンパク質の熱安定性の評価も同時に行うことができます。
タンパク質フォーカストライブラリー
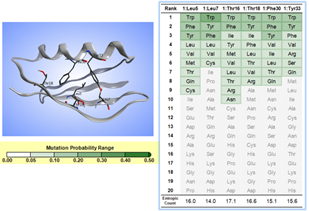 バーチャルファージディスプレイにより、リガンドとの親和力や熱安定性の向上、目的の物性値を持つタンパク質候補を効率的に構築することができます。ランダムサンプリングにより構築させた変異体構造において各種スコアや物性値を計算し、それぞれのポジション毎の活性体に含まれるアミノ酸出現頻度と情報エントロピーを算出します。その結果をもとに、膨大な組み合わせの中から目的の物性にフォーカスしたライブラリーに絞り構築します。
バーチャルファージディスプレイにより、リガンドとの親和力や熱安定性の向上、目的の物性値を持つタンパク質候補を効率的に構築することができます。ランダムサンプリングにより構築させた変異体構造において各種スコアや物性値を計算し、それぞれのポジション毎の活性体に含まれるアミノ酸出現頻度と情報エントロピーを算出します。その結果をもとに、膨大な組み合わせの中から目的の物性にフォーカスしたライブラリーに絞り構築します。
タンパク質表面パッチ解析
疎水性、正電荷、負電荷の傾向が強いタンパク質表面の一部(表面パッチ)を可視化します。表面パッチは、タンパク質の凝集やタンパク質間相互作用、溶解度などに影響します。凝集を防ぎ溶解度を向上するための残基の特定や、PPI阻害剤のターゲットの探索に活用できます。表面パッチの二次元への投影図は、野生型と変異体との比較などに利用できます。
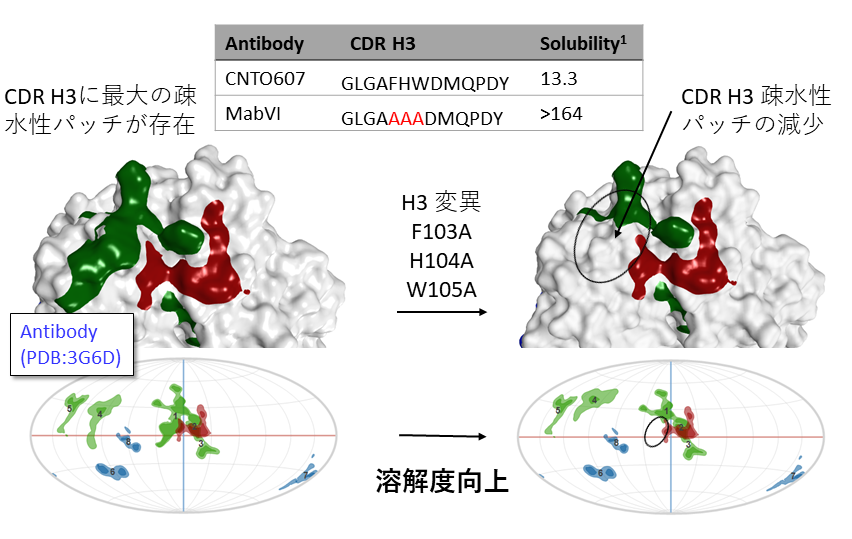
抗体CNTO607(左図)は高濃度で凝集。CDR H3にある疎水性パッチが多量体形成の原因となりうる。疎水性パッチ領域にアラニンを導入した変異体(右図)は疎水性パッチが減少し溶解度の向上が見られた。
1. Wu, S.-J.; et al. Protein. Eng. Des. Sel. 2010, 23 (8), 643–651.
タンパク質(核酸)-タンパク質ドッキング
タンパク質-タンパク質あるいは核酸-タンパク質間のドッキングシミュレーションは、単体のタンパク質、核酸の構造から、それらが形成する複合体の構造を予測します。得られた複合体の構造はタンパク質の重要部位の特定(エピトープマッピング)、タンパク質間の相互作用の解析・設計に利用できます。計算は粗視化モデルとグリッドを用いて高速化しています。探索範囲を抗体のCDRや酵素活性部位などに限定して計算を行うことができます。
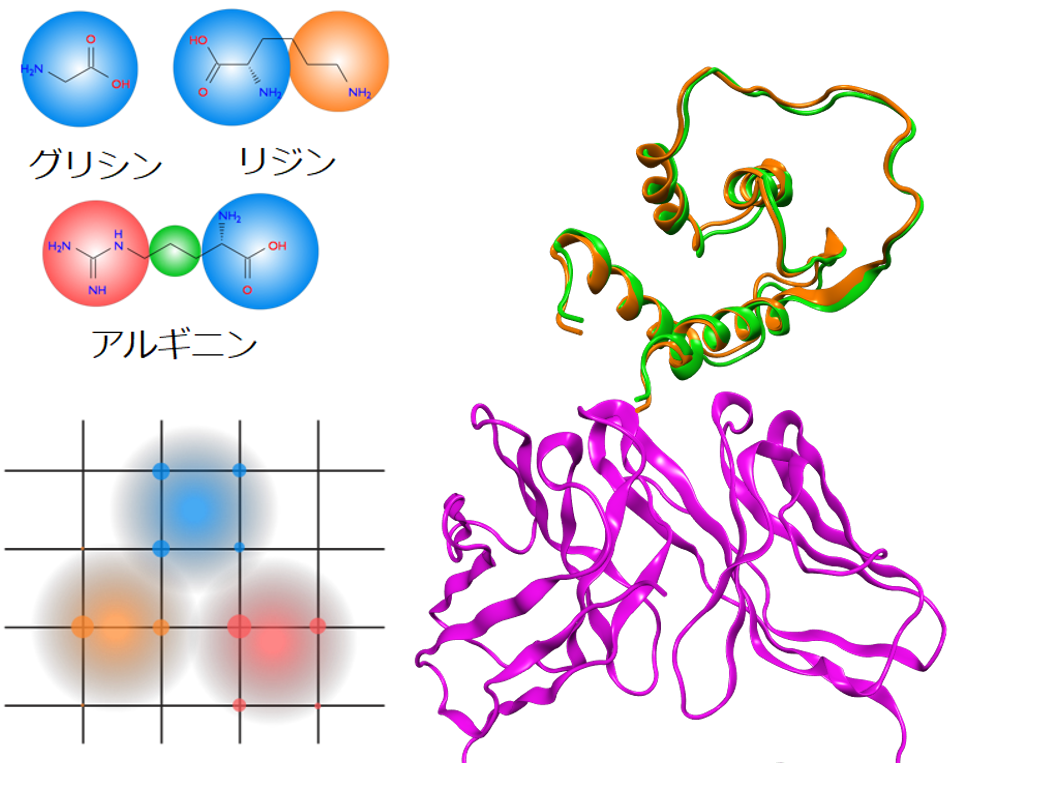
アミノ酸残基の粗視化イメージ(左上)、リガンドのガウス関数で重み付けされた密度のイメージ(左下)、抗体(右下、紫)とインターロイキン13(右上、実験結果:橙、計算結果:緑)のドッキング構造(PDB ID: 3G6D、RMSD: 0.8Å)
ループグラフティング
タンパク質の任意の部分構造をほかの構造より移植したループグラフティングが可能です。
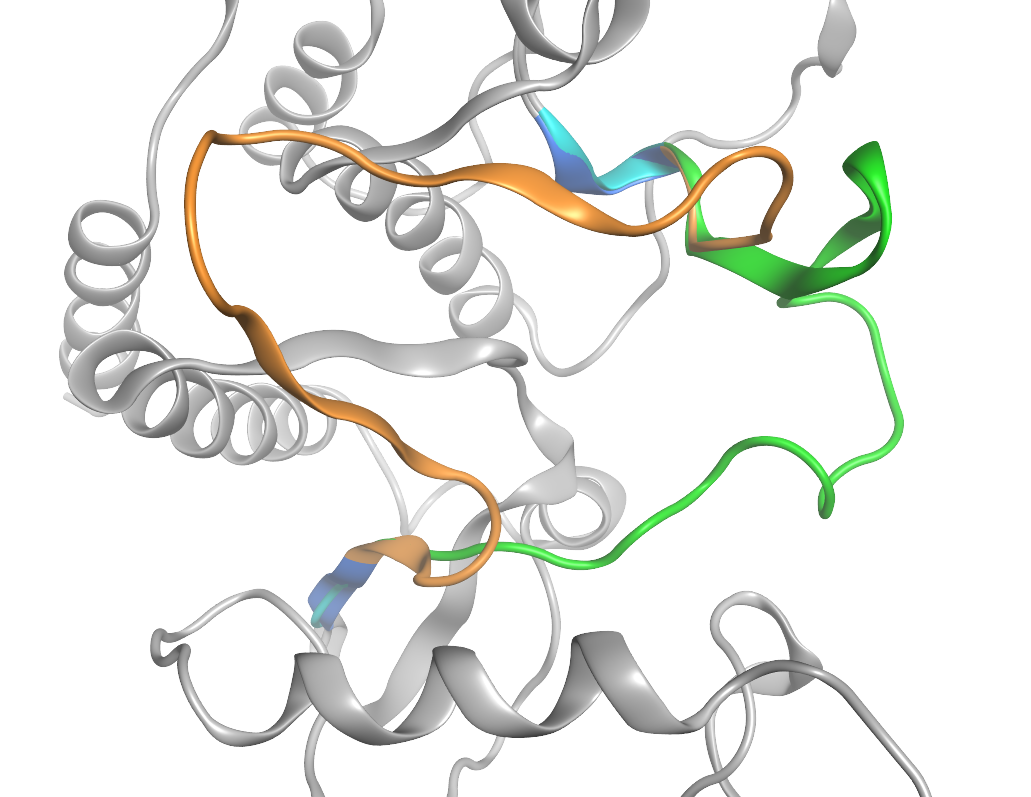
c-Ablキナーゼの活性化ループのループグラフティングのプレビュー表示(橙:移植前のループ、緑:移植後のループ、青:移植先の隣接残基、水色:移植元の隣接残基)
ループ/リンカーモデリング
ループ/リンカーモデリングは、タンパク質のループ構造を対話的に構築する機能です。ループ構造の構築手法として、PDBデータ内から検索する経験的探索と、新規のループ構造を構築するde novo探索の二種類が提供されています。溶媒和を考慮した相互作用エネルギーや、リガンドとの親和性、電子密度マップとの適合性を評価に加えることができます。リンカーモデリングでは、2つのタンパク質を融合するためのリンカー候補を探索します。この機能は、SBDD分野におけるタンパク質のループ構造のサンプリング、CABD (Computer-Aided Biologics Design)分野における融合タンパク質の構築、結晶解析分野における電子密度によるループ構造の補完など幅広い分野に活用できます。
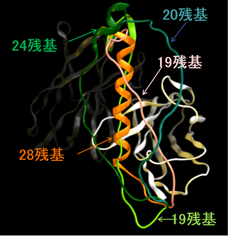
VLとVHと一本鎖抗体の構築例(PDB ID:1VFB)。
エピトープマッピング
エピトープマッピングは抗原の抗体結合部位(エピトープ)を同定することです。同定されたエピトープの情報は、新たな抗体医薬を設計する上で大変有用です。MOEのエピトープマッピングの機能では、タンパク質-タンパク質ドッキングの結果に対して、表面パッチを取り入れた相互作用フィンガープリントを用いたクラスタリングと相互作用エネルギーによるランク付けを行って、エピトープのアミノ酸残基を予測します。
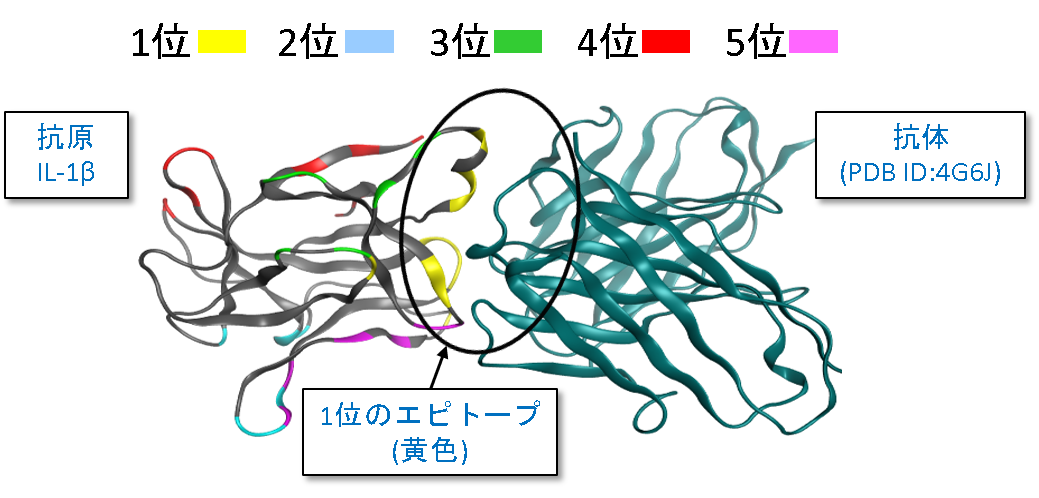
インターロイキン-1β(IL-1β)の上位5件のエピトープ予測例
タンパク質の相互作用解析
タンパク質の相互作用解析機能では、水素結合、イオン相互作用、vdW相互作用、共有結合、CH…O相互作用、π…H相互作用、ハロゲン結合、配位結合、アレーン接触を検出できます。分子内相互作用やタンパク質間相互作用(Protein-Protein Interaction)を解析できます。残基番号を軸の値に利用した2次元グラフを表示できます。データ点を選択すると、対応する3次元構造を選択・可視化が容易にできます。
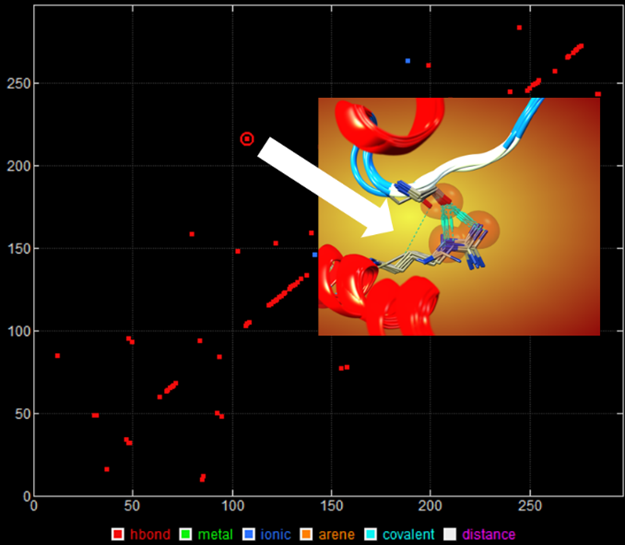
上皮成長因子受容体で保存された分子内相互作用の2次元グラフ(左)、選択データ点の3次元構造(右)
タンパク質物性推算/タンパク質記述子

タンパク質記述子の例
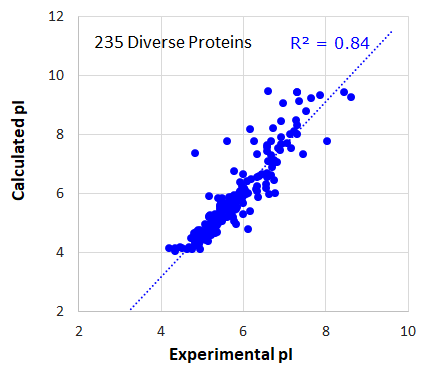
235個の様々なタンパク質構造における等電点(pI)の実験値と予測値の相関図
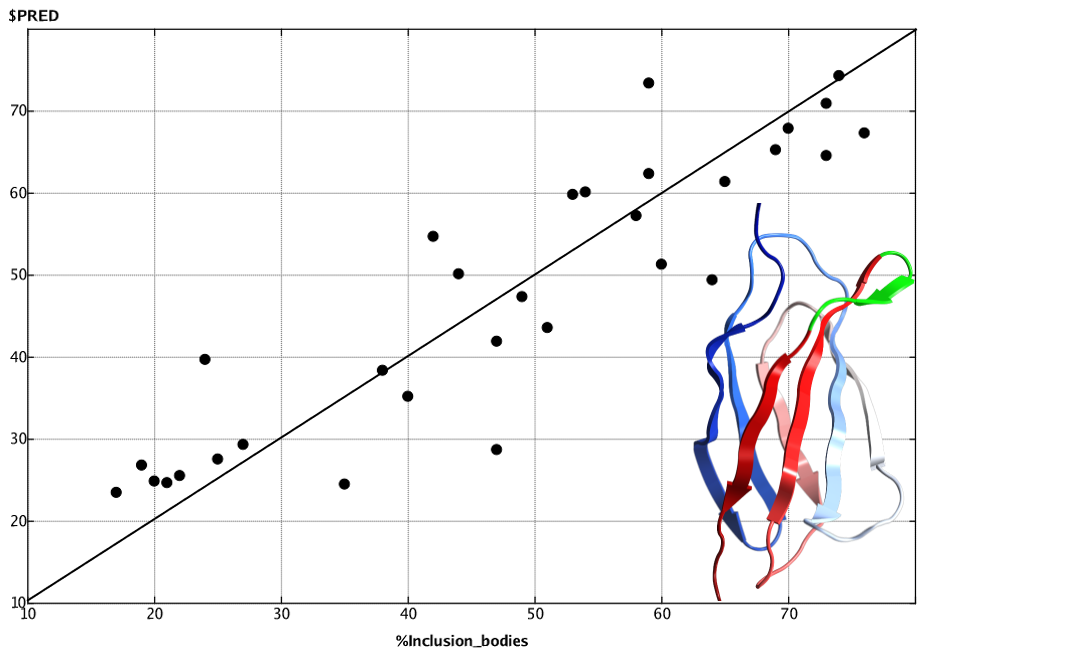
FGループの配列が異なる31個のアドネクチンの溶解度の予測モデル(決定係数:0.8183、Leave One Outによる交差検定の決定係数:0.7570)。縦軸が予測値、横軸が実験値。予測モデルには、疎水性モーメント、疎水性表面パッチ、正電荷表面パッチに関する4個のタンパク質記述子を使用。
ドメインモチーフ検索
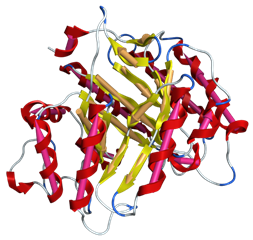 配列非依存的なタンパク質の類似構造検索が行えます。標準搭載のタンパク質立体構造データベースや、任意のタンパク質立体構造セットに対して、二次構造ベクトルの集合をモチーフ構造として検索します。この手法により、既存の検索法では検出できない遠縁のタンパク質も検出できます。また、ポケット類似構造検索などにも利用可能です。
配列非依存的なタンパク質の類似構造検索が行えます。標準搭載のタンパク質立体構造データベースや、任意のタンパク質立体構造セットに対して、二次構造ベクトルの集合をモチーフ構造として検索します。この手法により、既存の検索法では検出できない遠縁のタンパク質も検出できます。また、ポケット類似構造検索などにも利用可能です。
- PSILO
- タンパク質立体構造情報データベースシステム
- Daylight
- 情報化学システム構築ツール
- Chemotargets CLARITY
- ターゲット・作用機序予測プラットフォーム